El músculo esquelético es de contracción voluntaria, a diferencia del músculo liso, que es de contracción involuntaria. El músculo necesita esta energía para ejercer esta función contráctil. Dicha energía se obtiene a partir del ATP proveniente de la oxidación más o menos rápida de diferentes sustratos. Sin embargo, el músculo esquelético dispone de un reservorio de creatina, que una vez transformada en creatina-fosfato, proporciona energía de forma inmediata para la contracción muscular.
ANATOMÍA DE LOS MÚSCULOS
Las células del músculo esquelético, también llamadas fibras musculares, porque su aspecto es alargado (pueden alcanzar varios cm), y son el resultado de la fusión de varias células. Las fibras se agrupan en haces que permanecen unidos por una membrana de tejido conectivo. A su vez, varios haces se agrupan y son rodeados por una capa de tejido conectivo llamada epimisio. Todo este conjunto es lo que se conoce como músculo.
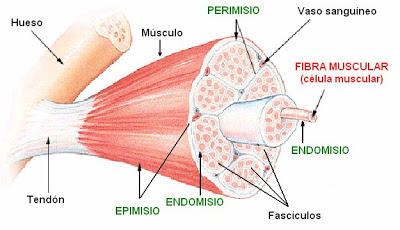
Anatomía de los músculos
El aparato contráctil muscular (miofibrillas) se pueden ver con la observación microscópica de un corte longitudinal de tejido muscular. En este corte se puede observar una sucesión ordenada de bandas trasversales. En este sentido se pueden distinguir:
- Bandas A: anisótropas (no dejan pasar la luz) y oscuras. Éstas presentan una zona clara denominada zona H, con una línea central denominada línea M.
- Bandas I: isótropas (dejan pasar la luz). Presentan una línea oscura denominada línea Z.
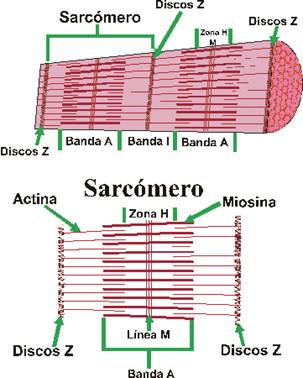
Bandas y líneas de los músculos
Estas bandas transversales son distintas zonas de apilamiento de microfilamentos longitudinales. Estos microfilamentos forman la base molecular del sistema contráctil de la fibra muscular. La unidad contráctil o sarcómero es la estructura comprendida entre 2 líneas Z. Allí se distinguen 2 tipos de filamentos de naturaleza proteica: filamentos gruesos de miosina y filamentos delgados de actina. Ambos filamentos se solapan en algunas zonas formando las bandas antes mencionadas. Cada filamento grueso está formado por 400 moléculas de miosina. Cada molécula de miosina es una doble hélice de 2 cadenas proteicas. La cabeza de las cadenas posee dominios moleculares de interacción con actina y de actividad ATPasa. Las cabezas sobresalen de la estructura y conectan con los filamentos delgados en 6 direcciones diferentes. Cada filamento delgado es una doble hélice, pero cada hebra es un rosario de monómeros de actina. La polimerización se realiza en presencia de ATP, magnesio y calcio. El doble rosario de actina se retuerce alrededor de una proteína con estructura de hélice alfa denominada tropomiosina. Finalmente aparece una proteína globular denominada troponina, que se sitúa a intervalos regulares sobre la actina.
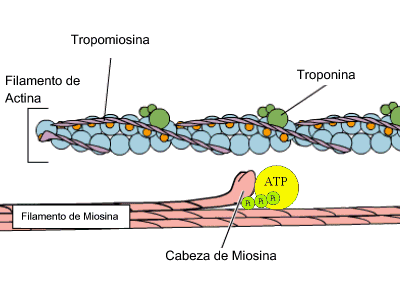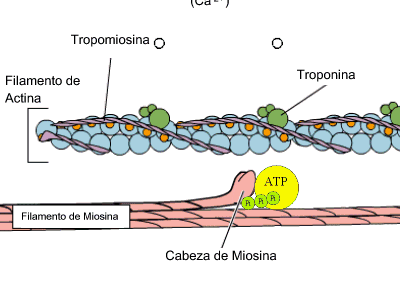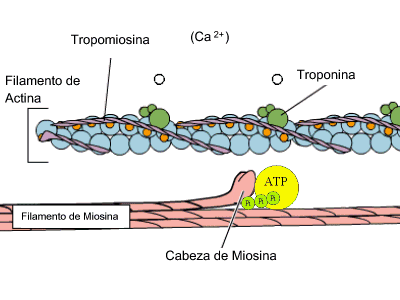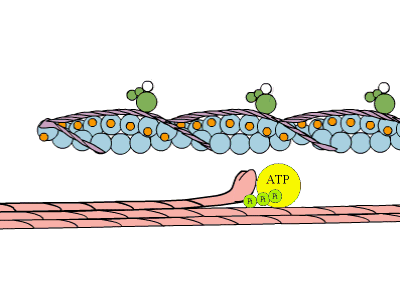
Actina y miosina, contracción muscular (GIF)
MECANISMO DE LA CONTRACCIÓN MUSCULAR
La estimulación muscular es desencadenada por un impulso nervioso, que produce una despolarización de la membrana celular y con ello la salida de iones de calcio del retículo sarcoplásmico. La concentración de calcio se incremente en un periodo muy corto de tiempo (ms). Los iones de calcio se unen a la troponina induciendo un cambio conformacional, que produce un desplazamiento de esta proteína con respecto a las moléculas de actina. Este desplazamiento hace posible el contacto de las moléculas de actina con las cabezas de miosina de los filamentos gruesos. En dichas cabezas se produce a la vez la hidrólisis de ATP rindiendo ADP y piruvato. Esto provoca un cambio conformacional de la cabeza que conlleva el desplazamiento del filamento delgado con respecto al grueso, el acercamiento de las líneas Z y por consiguiente la contracción muscular. Este proceso se repite hasta que la concentración de calcio disminuye por cese del estímulo nervioso.
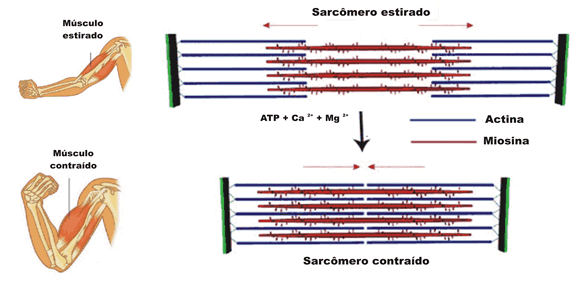
Contracción muscular
Para este proceso el músculo necesita mucho ATP, proporcionado por las mitocondrias a partir del ciclo de los ácidos tricarboxílicos en presencia de suficiente oxígeno. En casos de estrés y de esfuerzos duros, la obtención de ATP transcurre por la vía glucolítica hasta el lactato.
METABOLISMO DE LOS MÚSCULOS
Desde un punto de vista metabólico, existen 2 tipos de fibras musculares:
- Las que predomina un metabolismo aeróbico (oxidación completa de los sustratos a CO2 y H2O), estas se conocen también como fibras de tipo I o fibras lentas. Son fibras con un alto contenido en mioglobina (la proteína que toma el O2 que lleva en sangre la hemoglobina y lo lleva al interior del músculo) y por ello, con una abundante red de capilares. En su interior predomina la presencia de mitocondrias, donde se oxidan los principales sustratos energéticos: hidratos de carbono y ácidos grasos. Dado que la oxidación mitocondrial lleva su tiempo, estos músculos permiten realizar tareas sostenidas en el tiempo de intensidad baja-media. Son los músculos que predominan en los deportistas que practican disciplinas aeróbicas extensivas, como maratones, ciclismo en ruta o triatlones entre otros.
- Por otro lado están las fibras en las que predomina el metabolismo anaeróbico (producción de lactato). Estas fibras se denominan de tipo II o fibras rápidas. Son fibras con menos mitocondrias que las de tipo I, en las que el glucógeno es degradado hasta piruvato, el cual, en lugar de tomar la ruta oxidativa mitocondrial, es transformado en lactato por la acción de la lactato deshidrogenasa. Estas fibras son muy eficientes en contracciones extremadamente rápidas, como una carrera de corta distancia o un lanzamiento. Estas fibras consumen los sustratos de forma muy rápida, obtienen menor rendimiento en producción de ATP y además, acidifican la fibra muscular al producir ácido láctico, lo cual disminuye la eficiencia de la contracción muscular. Hay que añadir a esto, el gasto energético que debe realizar el hígado para reciclar de nuevo este lactato, que es expulsado al torrente sanguíneo, en glucosa. No obstante, las personas tienen una mezcla de fibras en las que tiende a predominar un tipo sobre otro, regla que se cumple incluso dentro de un mismo músculo.
Respecto a la utilización metabólica de los sustratos, conviene señalar algunos apuntes en el metabolismo muscular. Por ejemplo, la glucosa es captada del torrente sanguíneo por un transportador dependiente de insulina, denominado GLUT-4. Esto permite que cuando hay abundancia de sustrato (glucosa), lo que implica presencia de insulina en sangre, se pueda captar glucosa a nivel muscular para ser almacenada en forma de glucógeno o para ser utilizada en la vía glucolítica. Todo ello depende de si la persona está en reposo o está haciendo alguna actividad.
Los ácidos grasos también son un importante sustrato energético, sobre todo en fibras de tipo I. Estos ácidos grasos pueden tener varios orígenes. Pueden venir de las reservas de triglicéridos instramusculares, pueden venir por la circulación del tejido adiposo unidos a la albúmina de forma no esterificada o pueden venir de la dieta, o tras pasar por el hígado, esterificados en forma de lipoproteínas. Los ácidos grasos no esterificados pueden ser utilizados de forma más rápida por el músculo.
La utilización de ácidos grasos se ve favorecida por el ejercicio de intensidad baja-media, pero también en reposo o en situaciones de ayuno prolongado.
BIBLIOGRAFÍA
- Apuntes Universidad.